Eduard Strasburger, eine zentrale Figur in der Geschichte der Botanik, widmete einen erheblichen Teil seiner wissenschaftlichen Arbeit der Untersuchung von Befruchtungsvorgängen bei verschiedenen Pflanzenarten. Seine Forschungen, die sich auf Moose, Farne und Nacktsamer konzentrierten, legten wichtige Grundlagen für das Verständnis der zellulären Prozesse, die der Fortpflanzung zugrunde liegen.
Die Vererbung und damit die Kontinuität der Organismen in der Evolution beruhen nach dem Modell von August Weismann auf der "Kontinuität des Keimplasmas". Theodor Boveri leistete 1902 mit seiner Arbeit "Über mehrpolige Mitosen als Mittel zur Analys des Zellkerns" wichtige Beiträge zur Zellkernforschung.
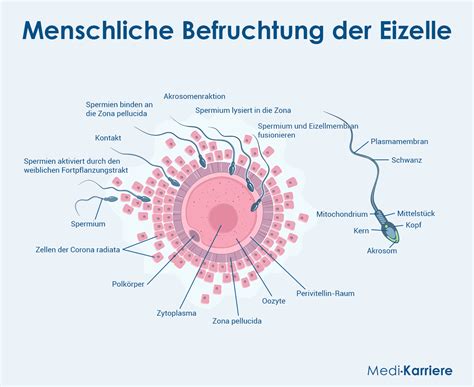
Die Rolle von Samenfäden bei der Befruchtung
Die Samenfäden gelangen in den Kanal des Archegonienhalses, durchlaufen diesen und treten schließlich ins Innere des Embryosacks ein. Sie durchbohren dabei die erweichte Membran der Scheitelregion des Embryosacks. Hier bewegen sie sich noch einige Zeit, wobei sie das nahe ihrer Eintrittsstelle an der Innenwand des Embryosacks angeschmiegte Keimbläschen munter umspielen. Unmittelbar nach Ankunft von Samenfäden wurde eine Trübung der Befruchtungskugel beobachtet; dieselbe wird körnig, undurchsichtig, was weitere Vorgänge in ihr erschwert.
Nach Beobachtungen von Hofmeister ist die erste Scheidewand stark gegen die Fläche des Prothalliums geneigt. Die äußere der so entstandenen Zellen ist die größere und wird durch eine neue, entgegengesetzt geneigte Scheidewand, die auf die erste Scheidewand aufgesetzt ist, erneut geteilt. Die so entstandene zweiflächige Zellstruktur entwickelt sich weiter.
Die Spermien bewegen sich mitunter in einer Schleimhülle, die den Wandbelag des Kanals bildet und sich in das umgebende Wasser ergießt. Nach dieser Entleerung bleibt in der Centralzelle nur die nackte Befruchtungskugel zurück. Sie rundet sich ab und zeigt am Scheitel, dicht über dem Zellkern, in günstigen Fällen eine hellere Stelle, die als Empfängnissfleck interpretiert wird.
Die Bläschen, in denen sich die Spermien befinden, sind klebrig, und die Spermien bleiben mit ihnen nicht selten an fremden Körpern hängen. Dabei ziehen und zerren sie, um sich zu befreien, was ihnen oft gelingt. Das hinterste Ende des Spermiums zieht sich dabei oft zu einem langen Faden aus, der schließlich reißt. Die Bläschen quellen im Wasser sehr bedeutend und haben sich das...
Das Spiralband schnellt mit einem Ruck auseinander und eilt davon. Nur in ganz seltenen Fällen bleibt etwas von der Spezialmutterzelle an dem Ort zurück, wo das Spermium gelegen; ihre letzten Spuren werden vollständig gelöst in dem Augenblick, wo das Spermium davon eilt.
Entwicklung des Archegoniums und der jungen Pflanze
Hofmeister beschreibt die Teilung der Scheidewand in zwei ungleiche Hälften: eine innere größere, die zur Centralstelle des Archegoniums wird, und eine äußere, etwas kleinere, welche nach wiederholter Teilung den Halsteil des Archegoniums erzeugt.
Bevor dies alles geschieht, soll sich in der Centralzelle um deren primären Kern eine zarte sphärische Zelle bilden, welche die Mutterzelle der jungen Pflanze ist. Strasburgers Beobachtungen weichen in manchen Punkten von denen Hofmeisters ab und schließen sich eher an das an, was Pringsheim bei Salvinia gesehen hat. Nachdem die ursprünglich Mutterzelle sich in eine innere und äußere Zelle geteilt hat, entwickelt sich zunächst die äußere Zelle weiter.
Die Kerne sind meist größer als die entsprechenden Kerne in den Halszellen, werden aber bald wieder aufgelöst. Eine Streifung der Kanalzelle, wie sie Pringsheim für Salvinia angibt, war hier nicht zu sehen. Nur die obersten Halszellen teilen sich weiter, die unteren werden entsprechend zu Dauerzellen, und nur selten erfolgt auch noch eine Teilung in ihnen. Das Wachstum schreitet nicht gleichmäßig zu allen Seiten des Halses voran. Die dem Einschnitt des Prothalliums zugekehrten...
Die Teilungen erfolgen zu beiden Seiten des Halses, so dass es auf Querschnitten völlig den Anschein gewinnt, als wachse derselbe mit einer Scheitelzelle, durch abwechselnd geneigte Scheidewände. Ebenso folgt aus der obigen Schilderung wohl zur Genüge, dass es keine einfach vierreihigen Hälse ohne Kanalzelle gibt. Bilder, die zu einer solchen Annahme verleiten, erhält man nur dann, wenn bei Seitenansichten der Hals des Archegoniums so zu liegen kommt, dass eine seiner radialen Längswände senkrecht...
Strasburgers Beiträge zur Zellbiologie und Genetik
Eduard Strasburger war ein Pionier in der Erforschung der zellulären Prozesse, die der Vererbung zugrunde liegen. Er beobachtete, dass die Befruchtung zwei aufeinander folgende Vorgänge beinhaltet: die Verschmelzung der Keimzellen und die Vereinigung der Zellkerne. Diese Beobachtungen, die er 1877 auf einem Botanikkongress in Amsterdam vorstellte, stießen zunächst auf heftigen Widerstand, da die Kernteilung wegen neuer Färbemethoden als Artefakt galt. Strasburger konnte jedoch nachweisen, dass Tochterkerne nur aus der Teilung von Mutterkernen hervorgehen können.
Er war maßgeblich an der Entwicklung des botanischen Lehrbuchs beteiligt, das heute noch verlegt wird. Die erste Auflage, die 1894 von Strasburger und drei Privatdozenten verfasst wurde, bot 558 Seiten Forschungsergebnisse und wurde in acht andere Sprachen übersetzt. Die aktualisierte 35. Auflage des Lehrbuchs verbindet bis heute ein Höchstmaß an Kontinuität mit einem Maximum an Aktualität.
Strasburgers Interesse galt Befruchtungsvorgängen in Farnen, Coniferen und bei Marchantia. Sein Postulat, die weiblichen Zapfen der Nadelhölzer seien kein Blüte, sondern ein Blütenstand, wurde erst 60 Jahre später durch einen Fossilienfund bewiesen. Er beschränkte sich nicht nur auf die Cytologie, sondern erforschte auch physiologische Prozesse.
Die Bedeutung Strasburgers für die Botanik zeigt sich auch an den von ihm geprägten Fachtermini wie Phototaxis, Zellplatte, Gamet, Cyto- und Nukleoplasma, Pro-, Meta- und Anaphase, Plasmodesmen, haploid und diploid. Fünf Universitäten verliehen ihm die Ehrendoktorwürde, und über 40 nationale und internationale Akademien, Gesellschaften und Vereine trugen ihm eine Mitgliedschaft an.
Moderne Forschung zur doppelten Befruchtung
Blütenpflanzen verfügen über einen komplizierten Befruchtungsmechanismus, die sogenannte doppelte Befruchtung. Hierbei entstehen ein Embryo und ein Nährgewebe, das Endosperm. Bei diesem Prozess verschmelzen jeweils zwei weibliche und zwei männliche Keimzellen, was zu zwei Befruchtungsprodukten führt: Aus dem ersten entwickelt sich der Embryo, der zur Pflanze heranwächst, und aus dem zweiten geht das Endosperm hervor, das als Nahrungsquelle für den Embryo dient.
Forscher haben jedoch herausgefunden, dass die doppelte Befruchtung auch umgangen werden kann. Durch experimentelle Ansätze gelang es, nach einfacher Befruchtung einen lebensfähigen Embryo zu erhalten, der während seiner Reifung von einem Endosperm ernährt wird, welches rein mütterlichen Ursprungs ist. Diese Erkenntnisse konnten eine über 100 Jahre alte Hypothese von Eduard Strasburger bestätigen.
Die Chromosomentheorie der Vererbung, die besagt, dass sich die materiellen Träger der Vererbung im Zellkern befinden, wurde durch die Arbeiten von Oscar Hertwig und Eduard Strasburger (1875-1877) maßgeblich gestützt. Heinrich Wilhelm Waldeyer führte 1888 die Bezeichnung Chromosomen ein. Walther Flemming beschrieb 1882 das Prinzip der Konstanz der Kernkörperchenzahl bei einer Zellteilung (Mitose), und Theodor Boveri beobachtete die Halbierung der Zahl der Kernkörperchen bei der Entstehung der Keimzellen.
Die Forschung zeigt, dass bei Pflanzen das Imprinting, eine Form der epigenetischen Regulation, bei der bestimmte Gene unterschiedlich stark exprimiert werden je nach elterlicher Herkunft, nicht zwingend notwendig ist, wenn das mütterliche und das väterliche Genom neutralisiert werden. Dies wirft Fragen nach der Rolle der männlichen Keimzelle bei der Entwicklung des Endosperms auf, wenn eine einfache Befruchtung ausreicht, um einen gesunden Embryo zu bilden.
Entwicklung des Embryosacks und Pollens (Gametophyten) bei Blütenpflanzen
tags: #eduart #strassburger #erkennt #befruchtung #geschlechtszellen